




APA
ISO 690-2
Harvard
Haga clic en un formato de citación
Evaluation of the Production of a Biocomposite Material from a Lignocellulosic Waste Mixture using Mycelium of Pleurotus ostreatus *
Evaluación de la producción de un material biocompuesto a partir de una mezcla de residuos lignocelulósicos utilizando micelio de Pleurotus ostreatus
Adriana Suesca Díaz ![]() , William Santiago Pedraza Baquero
, William Santiago Pedraza Baquero ![]() , Yovanny Morales Hernández
, Yovanny Morales Hernández ![]()
Evaluation of the Production of a Biocomposite Material from a Lignocellulosic Waste Mixture using Mycelium of Pleurotus ostreatus *
Ingeniería y Universidad, vol. 29, 2025
Pontificia Universidad Javeriana
Adriana Suesca Díaz a adriana.suesca@uamerica.edu.co
Universidad de América, Colombia
William Santiago Pedraza Baquero
Universidad de América, Colombia
Yovanny Morales Hernández
Universidad de América, Colombia
Received: 22 july 2024
Accepted: 03 june 2025
Published: 05 august 2025
Abstract: This study investigated the fabrication of a biobased material utilizing the Pleurotus ostreatus strain, with malt dust, coffee husks, and sugarcane bagasse serving as substrates. Six substrate mixtures were formulated and subjected to solid-state fermentation. The most effective formulation—comprising 50% sugarcane bagasse and 50% malt dust—achieved complete (100%) surface colonization. After 25 days of fermentation, significant increases of 54.5% in soluble lignin and 123.2% in protein concentrations were observed, accompanied by an 11.3% decrease in pH and a reduction of 0.118 g/mL in reducing sugars. These changes suggest strong metabolic interactions between the microorganism and agro-industrial residues, indicating efficient nutrient assimilation and promoting the formation of aerial mycelia in a reticulated structure, a key feature for the development of lightweight and insulating biomaterials. Although the biomaterial exhibited an average compressive stress of 255.5 ± 27.9 kPa, it has not yet reached the mechanical performance of EPS. However, these results represent a promising step forward in the development of sustainable materials.
Keywords:Chemical Kinetics, Mycelium, Pleurotus Ostreatus , Biocomposite Material, Lignocellulosic Waste, Solid Fermentation.
Resumen: Este estudio investiga la fabricación de un material biobasado utilizando la cepa Pleurotus ostreatus, empleando polvo de malta, cascarilla de café y bagazo de caña de azúcar como sustratos. Se formularon seis mezclas de sustratos y se sometieron a fermentación en estado sólido. La formulación más eficaz —compuesta por 50 % de bagazo de caña de azúcar y 50 % de polvo de malta— logró una colonización superficial completa (100 %). Después de 25 días de fermentación, se observó un aumento significativo del 54.5 % en la lignina soluble y del 123.2 % en las concentraciones de proteína, acompañado por una disminución del 11.3 % en el pH y una reducción de 0.118 g/mL en los azúcares reductores. Estos cambios sugieren una fuerte interacción metabólica entre el microorganismo y los residuos agroindustriales, lo que indica una asimilación eficiente de nutrientes y promueve la formación de micelio aéreo en una estructura reticulada, una característica clave para el desarrollo de biomateriales ligeros y aislantes. Aunque el biomaterial exhibió una resistencia a la compresión promedio de 255.5 ± 27.9 kPa, aún no alcanza el rendimiento mecánico del EPS. Sin embargo, estos resultados representan un paso prometedor en el desarrollo de materiales sostenibles.
Palabras clave: cinética química, micelio, Pleurotus ostreatus , material biocompuesto, residuos lignocelulósicos, fermentación sólida.
Introduction
The packaging industry extensively uses EPS due to its flexibility, light weight, and excellent compressive strength. However, its widespread use has led to significant environmental challenges because EPS is not biodegradable and contributes to the accumulation of plastic waste, known as white pollution. Recycling methods for EPS, such as mechanical operations, high melting temperatures, chemical recycling, and incineration, are currently employed. Despite their effectiveness, these methods are often expensive, produce toxic emissions, and involve hazardous solvents [1]. The annual consumption of EPS in Colombia is approximately 80,000 tons, with nearly 38% used for packaging and the remainder for industrial purposes. Despite this extensive use, the EPS recovery rate is low, with only 500 tons recycled annually. According to Garcia, this accounts for just 0.2% of the total plastic material recycled and a negligible 0.006% of the total EPS used [2]. The United States is the largest consumer, with a 50% share, whereas Europe and Japan divide the rest equally [3].
Recent advancements in biomaterials have propelled research into the creation of biodegradable materials through the colonization of fungi on organic substrates as an alternative to this problem [4]. This technique involves the rapid growth of mycelium, the vegetative part of the fungus, which produces many self-assembled linkages called hyphae, which are responsible for covering and degrading the lignocellulosic substrate, resulting in a natural and resistant material [4], [5]. After the fungus is deactivated and moisture is removed through heat, a biocomposite with properties similar to those of packaging materials is obtained [6]. These mycelium-derived biocomposites exhibit properties comparable to those of EPS, indicating their remarkable versatility in adapting to complex molds, biodegradability, and ability to be employed in applications such as thermal insulation, impact absorption, and cushioning [5].
Species such as P. pulmonarius and P. ostreatus are highly economically important for cultivation on lignocellulosic substrates [7]. P. ostreatus is widely used for its ability to produce ligninolytic enzymes, which mineralize lignocellulosic residues such as cellulose, hemicellulose, and lignin due to their strong oxidizing capacity [8], [9], [10]. These enzymes work by creating free radicals in biopolymers, destabilizing molecular chains, and breaking down macromolecules [10]. This enables the fungus to access a wide range of organic substrates for carbon and energy sources. Earlier studies have proven the feasibility of using fungal mycelia to produce biomaterials. In the research of [11], substrates such as sawdust, bagasse, and coffee husk, along with the P. ostreatus mushroom strain, were used to obtain a biomaterial with high density and compressive strength. Similarly, in the study of K. Joshi et al. [4], bioblocks with excellent physical and mechanical properties were manufactured from P. ostreatus mycelium grown on wastes, including wheat bran, sugarcane, sawdust, and mixtures of these substrates. Moreover, brewery waste, specifically barley dust, and the P. ostreatus strain were used to produce a biocomposite material with outstanding strength and elasticity properties [6]. These studies confirm that the properties of biomaterials made from mycelia are affected by numerous factors, such as the type of fungus used, the substrate used, the mycelial growth conditions, the manufacturing process, and the presence of additives [7].
In our previous research, we utilized various substrates to develop a solid fermentation The substrates used included the following residues: wheat bran [12], wheat bran and malt bran [13], and capacho of cape gooseberry with malt bran mixtures [14]. The C/N ratio is crucial for microorganism growth, and oxygen limitation in the substrates during solid fermentation hinders complete colonization of the residues. The present research shows the development and evaluation of the mycelial biocomposite material using malt dust, sugarcane bagasse, and coffee husk with P. ostreatus. The resulting material was physiochemically and mechanically characterized to find the most effective mixture for producing lignocellulosic-derived biomaterials with potential applications in industry.
Materials and Methods
Microorganisms
Pleurotus ostreatus was obtained from the ICA. To activate the microorganism, Sabouraud chloramphenicol agar was used. The mixture was incubated at 30±2 °C for 15 days according to Alemu et al. [11].
Substrates
Sugarcane bagasse (BG) was obtained as waste from panela production, and coffee husk (CH) was obtained as waste from the Cundinamarca region of Colombia. The substrates were predried by natural convection at 18 °C. Malt dust (MD) was obtained completely from the mill of a local craft brewery. Wheat bran was obtained from a commercial brand. The substrates were ground in a knife mill (grid diameter = 3 mm) and sieved below 1,70 mm. A 100 g sample was used for each substrate.
Solid-State Fermentation
Considering the abundance of sugarcane bagasse (BG) and coffee husk as agro-industrial waste, different substrate mixtures were proposed, and their theoretical C/N ratios were calculated (Table 1). The 25 gram mixtures were prepared in duplicate on polyethylene bags, presoaked with a 1:1 water ratio for 12 hours, and sterilized at 121 °C for 20 minutes [15]. The inoculum of P. ostreatus was cultivated on PDA plates at 30±2 °C for 7 days. The bags were aseptically inoculated with the mycelium obtained from the agar at an inoculum:substrate ratio of 1:4 (w/w). Finally, the samples were incubated for 19 days at 30±2 °C. After the incubation period, the materials were dried at 70 °C for 48 hours to stop microbial development, and the moisture in the biomaterials was extracted according to [16]. An observation of the relative mycelial colonization capacity was carried out, assigning 100% colonization to the positive control, which consisted of mycelial growth on wheat bran. From this reference point, the percentages of growth in the prepared mixtures were determined.
SSF Growth Monitoring Over TimeSSF Growth Monitoring Over Time
The lignocellulosic mixture with the highest mycelial growth observed qualitatively during solid-state fermentation was used. The bags, each with a total weight of 10 g, were prepared following the same procedure used for solid-state fermentation. The mixture was soaked in a 2:3 water-to-substrate ratio. The inoculated bags were sealed with a cotton and gauze plug and tightened with a 1/2-inch steel rod tube. Finally, the samples were incubated for 25 days at 30±2 °C in darkness. The mycelial growth rate was periodically inspected with photographic monitoring, moisture content, pH, total protein quantification, lignin measurement, and determination of reducing sugars to evaluate the colonization and growth of mycelial biomass in SSF over time or after 25 days of fermentation. Uninoculated bags were used as a negative control to compare the physicochemical characteristics of the substrate.
Physicochemical Characterization
The pH of fermentation was determined following the procedure proposed by Cujilema-Quitio et al. [21]. First, 1.0 g of dry biomass was suspended in 9.0 mL of distilled water. The sample was vortexed for 5 minutes and separated by vacuum filtration. The pH of the supernatant was subsequently read. Moisture determination was determined using the gravimetric method by subjecting the samples to drying in a forced convection oven at 65 °C for 24 hours[13]. Lignin determination of the samples was performed using acid hydrolysis with H.SO. solution (72% w/w) according to the NREL/TP-510-42618 procedure [22]. The filtered acid extract was analyzed at λ = 240 nm (bagasse) to measure the soluble lignin content that could have been released. For the determination of reducing sugars, the supernatant obtained during the pH measurement was used, and the dinitrosalicylic acid method was applied with a glucose standard solution[23]. For protein determination, the biuret method was employed, using the procedure described in [24], [25]. A calibration curve was prepared using a bovine serum albumin (BSA) standard solution. A total of 1.5 grams of sample was macerated with 20 mL of 50 mM PBS (pH 8) and shaken for 60 minutes at 180 RPM. After the mixture was vacuum filtered, the total protein content of the extract was determined by spectrophotometry at λ = 540 nm [26].
Compressive Strength
After 25 days of solid-state fermentation, fresh samples from 25-day bags were molded with sterile cylindrical molds, placed in sterile translucent PET bags and incubated at 30±2 °C in darkness for an additional 8 days to ensure the structural stability of the samples [13]. The molded biomaterials were then dried at 65 °C for 48 hours and stored in a vacuum desiccator. Compression tests were conducted using a Shimadzu AGS-50kNX universal testing machine, following ASTM D695-23 standards, on samples with an average diameter of 15.67 mm and a height of 15.43 mm at a speed of 1 mm/min.
Morphological Analysis of Mycelium Fibers
The dried samples were cut into small squares approximately 3 × 3 mm in size using a scalpel. The surface morphology of the mycelium fibers grown on the biomaterial was analyzed by scanning electron microscopy (SEM) (Zeiss evo HD 15) EHT using an accelerating voltage of 15.00 kV at the Engineer Faculty, Pontificia Universidad Javeriana, Bogotá, Colombia. The average diameter of the mycelium on the lignocellulosic substrate was calculated using ImageJ software.
Statistical Analysis
The data were statistically analyzed using ANOVA followed by Tukey's post hoc test, which was performed in RStudio v14.0, with a 95% confidence level.
Results
Substrates
The particle sizes of sugarcane bagasse and malt dust were more heterogeneous than that of coffee husk (Figure 1). Coffee husk had a more uniform distribution, with 70% of the particles at 1.70 mm. Malt dust had 56.2% of the particles between 0.25 mm and 0.85 mm, and sugarcane bagasse had 52.4% of the particles in that range and 24.4% between 0.60 mm and 0.71 mm. These results indicate that coffee husks were more uniform, whereas malt dust and sugarcane bagasse had greater variability in terms of particle size, indicating particle heterogeneity.
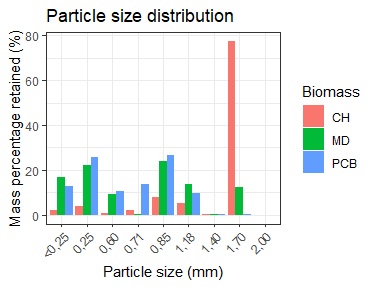
Notes. CH: Coffee husk, MD: Malt dust and PCB: Sugarcane bagasse.
According to Mercado et al., particle size significantly affects mycelial growth by influencing access to nutrients, air, and water. Large particles greater than 2 cm in size hinder substrate compaction and nutrient access, whereas small particles less than or equal to 2 cm limit aeration[26]. Presieving sugarcane bagasse (BG) can help to obtain particles smaller than 2 cm that support microbial growth and enhance compaction. Conversely, coffee husk (CH) has large particles, which are unfavorable for mycelium gas exchange due to their thickness and fineness, allowing the substrate to slide through mill blades without breaking.
Solid Fermentation
After 19 days of incubation, the mycelium covered the surface mixture in the control experiment. The mixture M. (sugarcane bagasse–malt dust 50:50) (Figure 2) achieved the highest percentage of colonization by the fungus (i.e., between 90% and 95%), with no significant differences compared with the control (Figure 3), followed by mixture M. (coffee husk–sugarcane bagasse 50:50), with approximately 85–90%, and M. (coffee husk–sugarcane bagasse 80:20), with 80–85% colonization. M1 and M2 sugarcane bagasse 100% and sugarcane bagasse–malt dust 80:20 had colonization percentages less than 25%, which were significantly different from those of the other mixtures. However, no significant differences were observed among mixtures M3, M5, and M6, suggesting that the inclusion of sugarcane bagasse in the formulations may be adequate to compensate for its low nitrogen content and improve the C/N ratio. The growth differences between the mixtures could be due to the variation in aeration between the substrate particles, the nutritional content, and the difficulty in extracting nutrients from compacted substrates [11]. With respect to mixture M., the higher growth rate of the mycelium on the bagasse substrate was attributed to the properties of its softer particles, which, according to [27], allow the fungus to extract nutrients more efficiently.

According to [28], the elemental composition of a substrate plays a fundamental role in the growth and development of fungal mycelia. In this sense, the mycelial growth rate in this study aligns with the findings of [16], where complete colonization of P. ostreatus mycelium on sugarcane bagasse, coffee husk, sawdust, and chopped dry grass supplemented with construction gypsum and rice flour occurred in approximately 18 days. This is supported by the research of [29], where a 50% increase in P. ostreatus production was observed when sunflower husk flour was used as a nitrogen supplement. According to the authors, enriching substrates with nitrogen is also a crucial factor in increasing the protein content of fungi.
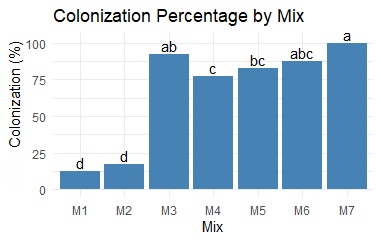
Additionally, the results are compared with those of [30], who reported that the growth time of P. ostreatus was prolonged when the proportion of nitrogen in the substrate was lower. Their study [30] revealed that the fungus P. ostreatus, which is responsible for the decomposition of lignocellulosic substrates, is strongly dependent on low C/N ratios. Despite the favorable mycelial development observed in the mixtures containing coffee husks, these mixtures were discarded because of their fragility and susceptibility to fracture after drying. This could be linked to the large particle sizes impeding nutrient access. In contrast, the M3 mixture showed significantly high colonization and a particular resistance to manual compression. The heterogeneity of sugarcane bagasse could aid in mycelium adaptation and nutrient absorption, promoting biomass formation.
SSF Growth Monitoring Over Time
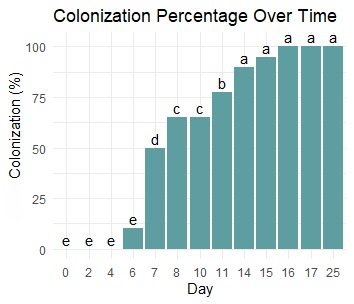
A second solid-state fermentation (SSF) using a mix of M3 (sugarcane bagasse–malt dust 50:50) was developed (Figure 4). Mycelial growth and 100% growth were obtained on Day 16 of experimentation. No statistically significant differences were found among the samples between Days 14 and 25 of fermentation (Figure 5). Using a cotton and gauze plug with a steel rod tube improved gas transfer and decreased the degree of oxygen limitation, decreasing the colonization from 16 days. This method of SSF increased cellular respiration and led to more uniform mycelial growth and faster colonization. On the 25th day of experimentation, the mycelium not only colonized the substrate but also began to develop a greater amount of aerial mycelium that was distributed along the bag in an upward direction, indicating the availability of nutrients for continued development. These results demonstrate the direct consumption of nutrients provided by the substrates, especially by sugarcane bagasse, due to its high nutrient content [4]. The estimated percentage of colonization is shown in Figure 5, which was determined by observing the surface of the bags, where the beginning of colonization occurred on Day 6. Notably, this rapid colonization continued with the appearance of dense, white, and extensively branched mycelia that completely enveloped the substrates inside the bags after 14 days of growth.

Physicochemical Characterization
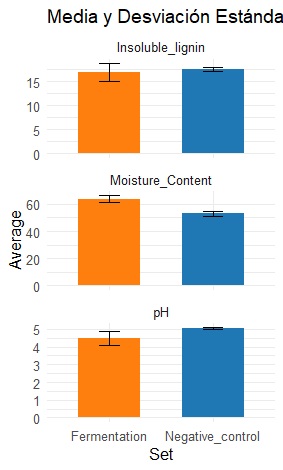
The pH of the negative control mixture was 5,04 ± 0.03; in contrast, the final pH of the replicates of the inoculated mixtures was 4,47 ± 0,19. There was a statistically significant decrease in pH during fermentation (Figure 6). This process is associated with the secretion of enzymes by the fungus [9], [31]. These enzymes breakdown components such as cellulose, hemicellulose, and lignin in the substrate, releasing organic acids such as oxalic acid and formic acid [32], which in turn cause the observed decrease in pH. In this way, the findings of [33], [34] suggest that the decrease in pH due to the enzymatic digestion process is a result of mycelium colonization.
The moisture content of the samples at the end of fermentation (63,32 ± 1,618%) was significantly greater than that of the negative control samples (52,91 ± 2,671%) (p < 0,05) (Figure 6). This finding indicates that part of the metabolic water was contained inside the bags during fermentation as the mycelia developed. This phenomenon, attributed to the enzymatic digestion process of the mycelium, was described by Attias et al. [27]. The results of moisture tests revealed that the percentages of mycelial growth in SSF in the bags are within the optimal percentages, with values ranging between 62% and 65% [24].
The insoluble lignin content of the samples at the end of fermentation was 16,90 ± 0,78%, and that of the control was 17,51 ± 0,59% (p > 0,05); because these values were not significantly different (Figure 6), it was not possible to determine an insoluble lignin decrease due to fungal growth. However, an increase in soluble lignin levels was observed as fermentation time increased (Figure 7), with values ranging from 6,96–9,46% from 6–25 days. The statistical analysis revealed significant differences in the soluble lignin percentage across days. This result is favorable, as it highlights the enzymatic behavior of the P. ostreatus fungus. The extracellular enzymes secreted by fungi are essential for the initial transformation of lignin and its subsequent mineralization [24].
Note. Error bars represent the standard deviation.
The average protein content of the samples at the end of fermentation was 1.849 ± 0.001% mg/mL, whereas that of the control was 0.828 ± 0.0034 mg/mL (p > 0,05). Microbial proteins can be related to the consumption of free sugars present in the substrate, resulting from the possible enzymatic degradation of cellulose and hemicellulose. In terms of protein content, interference from substances present in the substrates can be observed. This is supported by the findings of [24], where the interference of the reported protein values was caused by the presence of starch and proteins in the substrate, which is one of the most common causes of data variation in solid-state fermentation. However, a statistically significant difference was observed between the protein concentration of the control and that measured over time (Figure 7), which may be associated with mycelial development. The protein concentration on Day 25 of fermentation was 123,2% greater than that of the negative control. These results are comparable to those reported in another study, where corn cob residues were used as substrates for the growth of P. pulmonarius [24], resulting in a 221.95% increase in protein content after 30 days of fermentation.
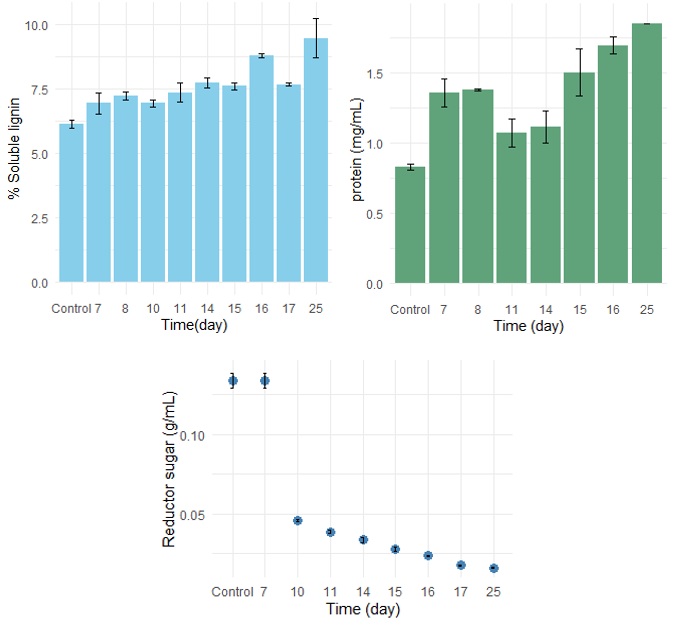
Note. Error bars represent the standard deviation.
The initial reducing sugar concentration during fermentation was 0,134 ± 0,006 g/mL on Day 6, followed by the gradual and significant consumption of these sugars throughout fermentation, reaching a minimum concentration of 0,016 ± 0,000 g/L on Day 25 (Figure 7). The enzymes of the microorganisms may breakdown the β-glucosidic bonds of cellulose in both its amorphous and crystalline regions [35]. Therefore, the release of sugar subunits during solid-state fermentation (SSF) facilitates enzymatic hydrolysis outside the lignocellulosic matrix, ultimately releasing glucose accessible to the fungus for microbial biomass synthesis [36]. This breakdown of cellulose and hemicellulose provides the fungus with an essential nutrient, glucose, which drives rapid mycelial growth [31].
Compressive Strength
The fungal biomaterial compressive strength was 255,5 ± 27,9 MPa, which was significantly greater than the value of 106 ± 1,12 MPa for EPS [37]. During the compressive strength test, the highest force supported was 50.02 kN, with a maximum deformation of 104.4%. Although the samples did not have a homogeneous surface, resulting in differences in cross-sectional area, similar values for deformation and maximum force were obtained. The materials tested showed a strain resistance of over 80% without fracturing. Compared with a biomaterial developed using 100% barley dust, as reported in [6], where the average stress for 10% deformation for the optimal formulation was 55,8 kPa, the values reported in this study are lower. However, previous studies using a 50:50 mixture of malt dust and wheat bran yielded a material with improved tensile strength of 110,04 kPa [13], demonstrating that the heterogeneity and nutritional composition of the initial substrate are key factors for homogeneous mycelial development and effective substrate utilization.
Morphological Analysis of Mycelium Fibers
The morphology of the hyphae of the fungus P. ostreatus grown on the lignocellulosic substrate was identified using SEM (Figure 8). Scanning electron microscopy (SEM) of sugarcane bagasse and malt powder without mycelium (control) (Figure 8a) revealed a rough and irregular surface as a result of the cutting treatment during milling.

Notes. a) Blank SEM image of M3 without colonization, 500X, scale bar 20 µm; b) M3 colonized by mycelium, 500X, scale bar 20 µm; c) M3 colonized by mycelium, 1,00KX, scale bar 10 µm; d) M3 colonized by mycelium, 2,50KX, scale bar 10 µm.
Likewise, the presence of small spheres, which were possibly malt dust, was observed. In contrast, the SEM images in Figure 8b show the colonization of P. ostreatus mycelia, which starts from the central part of the substrate and subsequently extends vertically upward. This growth pattern indicates that the mycelium takes advantage of the nutrients available in the substrate but also seeks oxygen in the surrounding atmosphere, which drives the formation of an ascending hyphal network. As shown in Figure 8d, the SEM micrograph of the mycelium grown on the lignocellulosic substrate reveals a dense network of typical P. ostreatus hyphae with tubular shapes that exhibit cuts during the process, anastomoses, septa, and clamp connections along their filaments. These observations are in line with the findings reported in a previous study [38]. Clamp connections, a hallmark of most Basidiomycota, arise during the cell division of secondary hyphae, whereas anastomoses facilitate hyphal fusion[11]. Previous studies have indicated that the diameter of a fully developed P. ostreatus mycelium is within the range of 1000–4000 nm [39]. The average diameter of the mycelium after 16 days of solid-state fermentation was calculated to be 2404 ± 15 nm. This study presents results on the morphological characteristics and diameters of mycelium fibers that are congruent and superior to the values reported in [4], where average diameter values of 2149 ± 111 nm were reported in sugarcane substrates and 2200 ± 81 nm in mixed media (wheat bran, sugarcane, and sawdust).
Conclusions
In conclusion, this study revealed that a mixture of agro-industrial waste composed of sugarcane bagasse and malt powder is highly favorable for the growth of P. ostreatus because of its high mycelial coverage, which provides sufficient nutrients for fungal growth. The growth of the microorganisms in mixtures of sugarcane bagasse, malt dust, and coffee husks was positive, achieving colonization levels above 75% after 19 days of fermentation. The M. mixture (sugarcane bagasse–malt dust 50:50) was chosen for monitoring over a 25-day fermentation, and correlations between the parameters analyzed were identified. The consumption of reducing sugars, the increases in protein and soluble lignin concentrations, and the decrease in pH were related to the increase in substrate colonization over the observed period of time. Visible mycelial development began on Day 6, with successful colonization achieved from Day 14 onward, indirectly indicating substrate consumption as a result of the fungus's metabolic activation. This process likely involves the production of lignin-degrading enzymes, reflected in the pH reduction, possibly due to the formation of organic acids. A net protein production of 1,340 mg/mL indicates positive fungal biomass synthesis. Significant consumption and release of reducing sugars were observed, with a net consumption of 117,936 mg/mL at 25 days. Changes in pH during fermentation indicated that the enzymatic activities of the fungus contributed to the consumption of lignocellulosic material, which was linked to microbial growth in solid fermentation.
The biocomposite material's mechanical compressive strength of 255.5 ± 27.9 MPa and strain resistance of over 80% support its potential as an alternative to traditional materials in the packaging industry, following a circular economy approach. However, additional mechanical tests and optimization of the mixed substrate are needed to confirm its use as a direct replacement for EPS. Overall, these findings could enhance the use of lignocellulosic waste to make sustainable materials.
References
[1] C. Sharma and K. Qanungo, “An overview: Recycling of expanded polystyrene foam,” AIP Conf Proc, vol. 2535, no. 1, p. 30019, May 2023, doi: 10.1063/5.0111982.
[2] N. García, “Evaluación del impacto ambiental de la aplicación de un plan de gestión posconsumo de poliestireno expandido (EPS) utilizado en el envase de alimentos en Colombia,” Thesis, Universidad EAN, Bogotá, 2019.
[3] J. Arandes, J. Bilbao, and D. López, “Reciclado de residuos plásticos,” Revista Iberoamericana de polímeros, vol. 5, no. 1, pp. 28–45, 2004.
[4] K. Joshi, M. K. Meher, and K. M. Poluri, “Fabrication and Characterization of Bioblocks from Agricultural Waste Using Fungal Mycelium for Renewable and Sustainable Applications,” ACS Appl Bio Mater, vol. 3, no. 4, pp. 1884–1892, Apr. 2020, doi: 10.1021/acsabm.9b01047.
[5] S. Sivaprasad, S. K. Byju, C. Prajith, J. Shaju, and C. R. Rejeesh, “Development of a novel mycelium bio-composite material to substitute for polystyrene in packaging applications,” Materials Today: Proceedings, vol. 47, no. 15, 2021. doi: 10.1016/j.matpr.2021.04.622.
[6] B. P. Bohórquez Sánchez and N. Ramírez Osma, “Desarrollo y caracterización de un material biodegradable a partir de desechos cerveceros y micelio del hongo Pleurotus ostreatus,” Thesis, Universidad de los Andes, Bogotá, 2020.
[7] A. Butu, S. Rodino, B. Miu, and M. Butu, “Mycelium-based materials for the ecodesign of bioeconomy,” Dig J Nanomater Biostruct, vol. 15, no. 4, pp. 1129–1140, 2020, doi: 10.15251/djnb.2020.154.1129.
[8] A. Doroški, A. Klaus, A. Režek Jambrak, and I. Djekic, “Food Waste Originated Material as an Alternative Substrate Used for the Cultivation of Oyster Mushroom (Pleurotus ostreatus): A Review,” Sustainability, vol. 14, no. 19, 2022, doi: 10.3390/su141912509.
[9] C. Sánchez, “Cultivation of Pleurotus ostreatus and other edible mushrooms,” Appl Microbiol Biotechnol, vol. 85, no. 5, pp. 1321–1337, 2010, doi: 10.1007/s00253-009-2343-7.
[10] J. Quintero, G. Feijoo, and J. Lema, “Producción de enzimas ligninolíticas con hongos basidiomicetos cultivados sobre materiales lignocelulósicos,” Vitae, vol. 13, no. 2, pp. 61–67, 2006.
[11] D. Alemu, M. Tafesse, and Y. Gudetta Deressa, “Production of Mycoblock from the Mycelium of the Fungus Pleurotus ostreatus for Use as Sustainable Construction Materials,” Advances in Materials Science and Engineering, vol. 2022, 2022, doi: 10.1155/2022/2876643.
[12] P. Leal Rocha and L. C. Rodriguez Moreno, “Desarrollo y evaluación de las propiedades mecánicas de un biomaterial para el desarrollo de biobloques obtenido a partir de biomasa fúngica y residuos agroindustriales,” Thesis, Fundación Universidad de América, Bogotá, 2022.
[13] A. Medina-Gutierrez, P. Medina-Rodriguez, A. Suesca-Díaz, and D. Morales-Fonseca, “Producción de un material biocompuesto a base de micelio por medio de fermentación sólida usando Pleurotus ostreatus,” Biotecnología en el Sector Agropecuario y Agroindustrial, vol. 22, no. 2, 2024.
[14] S. Castillo Larrarte and N. Lozano Escorcia, “Valorización de residuos de la industria cervecera artesanal empleando Pleurotus Ostreatus como estrategia de sostenibilidad ambiental para el sector,” Thesis, Universidad de América, Bogotá, 2024.
[15] L. S. Vásquez, V. Sopo, A. Suesca-Díaz, and D. Morales-Fonseca, “Elaboration of a Biomaterial from Pleurotus ostreatus Mycelium and Residual Biomass, as an Alternative to Synthetic Materials,” Chem Eng Trans, vol. 99, pp. 91–96, 2023, doi: 10.3303/CET2399016.
[16] E. Colmenares, L. Bautista, and C. Oliveros, “Material ecológico con fines de embalaje a partir del hongo Pleurotus ostreatus y residuos orgánicos agroindustriales,” Revista Científica Unet, vol. 30, 2018.
[17] M. Ruilova and A. Hernández, “Evaluación de residuos agrícolas para la producción del hongo Pleurotus ostreatus,” ICIDCA sobre los derivados de la caña de azúcar, vol. 48, no. 1, pp. 54–59, 2014.
[18] E. G. Silveira Junior, V. H. Perez, S. C. S. E. de Paula, T. da C. Silveira, F. L. Olivares, and O. R. Justo, “Coffee Husks Valorization for Levoglucosan Production and Other Pyrolytic Products through Thermochemical Conversion by Fast Pyrolysis,” Energies, vol. 16, no. 6, 2023, doi: 10.3390/en16062835.
[19] O. M. Menjura and Z. Sanchez, “Ajuste de un modelo matemático para la combustión de bagazo de caña en una cámara Ward-Cimpa,” Ciencia & Tecnología Agropecuaria, vol. 15, no. 2, pp. 133–151, 2014, doi: 10.21930/rcta.vol15_num2_art:355.
[20] J. Zhou, Y. Wu, D. Liu, and R. Lv, “The effect of carbon source and temperature on the formation and growth of Bacillus licheniformis and Bacillus cereus biofilms,” LWT, vol. 186, no. September, p. 115239, 2023, doi: 10.1016/j.lwt.2023.115239.
[21] M. C. Cujilema-Quitio, G. León-Revelo, M. Rizo Porro, L. Taramona Ruiz, and L. B. Ramos-Sánchez, “Producción de lipasas por fermentación sólida con Aspergillus niger: Influencia del pH,” Centro Azúcar, vol. 45, no. 4, pp. 1–9, 2018.
[22] Sluiter, A. D., & Templeton, D. W. “NREL/TP-510-42618 Determination of Structural Carbohydrates and Lignin in Biomass”, Technical report, NREL, 2008. http://www.nrel.gov/biomass/analytical_procedures.html
[23] G. L. Miller, “Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry, 31: 426-428.,” Analytical Chemistry, vol. 31, pp. 426-428, 1959.
[24] J. C. Oviedo, “Evaluación de la producción de proteína fúngica a partir de Pleurotus pulmonarius empleando residuos de poscosecha de maíz,” Thesis, Universidad de Antioquia, Medellín, 2014.
[25] Z. G. Li, Y. Q. Sun, W. L. Zheng, H. Teng, and Z. L. Xiu, “A novel and environment-friendly bioprocess of 1,3-propanediol fermentation integrated with aqueous two-phase extraction by ethanol/sodium carbonate system,” Biochem Eng J, vol. 80, pp. 68–75, 2013, doi: 10.1016/j.bej.2013.09.014.
[26] J. Mercado and J. Ttupa, “El bagazo de caña de azúcar como sustrato para la producción de Pleurotus ostreatus (hongo de ostra),” Thesis, Universidad Nacional de Callao, Callao 2023.
[27] N. Attias, E. Tarazi, and J. Y. Grobman, “Developing novel applications of mycelium based bio-composite materials for design and architecture,” Building with bio-based materials: Best practice and performance specification, vol. 1, no. 10, pp. 1-10, September, 2017.
[28] H. T. Hoa, C. L. Wang, and C. H. Wang, “The effects of different substrates on the growth, yield, and nutritional composition of two oyster mushrooms (Pleurotus ostreatus and Pleurotus cystidiosus),” Mycobiology, vol. 43, no. 4, pp. 423–434, 2015, doi: 10.5941/MYCO.2015.43.4.423.
[29] N. R. Curvetto, D. Figlas, R. Devalis, and S. Delmastro, “Growth and productivity of different Pleurotus ostreatus strains on sunflower seed hulls supplemented with N-NH4+ and/or Mn(II),” Bioresour Technol, vol. 84, no. 2, pp. 171–176, 2002, doi: 10.1016/S0960-8524(02)00013-5.
[30] M. B. Ruilova Cueva, A. Hernández, and Z. Niño-Ruiz, “Influence of C/N ratio on productivity and the protein contents of Pleurotus ostreatus grown in differents residue mixtures,” Rev. FCA UNCUYO, vol. 49, no. 2, pp. 331–334, 2017.
[31] D. Alemu, M. Tafesse, and A. K. Mondal, “Mycelium-Based Composite: The Future Sustainable Biomaterial,” International journal of biomaterials, vol. 1, 2022. doi: 10.1155/2022/8401528.
[32] M. Mäkelä, S. Galkin, A. Hatakka, and T. Lundell, “Production of organic acids and oxalate decarboxylase in lignin-degrading white rot fungi,” Enzyme and Microbial technology, vol. 30, no. 4, 2002, doi: 10.1016/S0141-0229(02)00012-1.
[33] N. Attias et al., “Mycelium bio-composites in industrial design and architecture: Comparative review and experimental analysis,” J Clean Prod, vol. 246, p. 119037, 2020, doi: 10.1016/j.jclepro.2019.119037.
[34] W. Aiduang, J. Kumla, S. Srinuanpan, W. Thamjaree, S. Lumyong, and N. Suwannarach, “Mechanical, Physical, and Chemical Properties of Mycelium-Based Composites Produced from Various Lignocellulosic Residues and Fungal Species,” Journal of Fungi, vol. 8, no. 11, Nov. 2022, doi: 10.3390/jof8111125.
[35] S. Montoya, “Obtención de enzimas lignocelulolíticas.” Thesis, Universidad de Caldas Manizales, 2012.
[36] K. S. Franco Peña, “Parámetros de crecimiento de Pleurotus ostreatus en cultivo semi-sólido,” Thesis, Universidad de los Andes, Bogotá, 2018.
[37] ASTM, “ASTM D695: Standard test method for compressive properties of rigid plastics,” ASTM International, vol. 08, April 2003, pp. 1–7, 2015, doi: 10.1520/D0695-23.2.
[38] I. Milovanović, I. Brčeski, M. Stajić, A. Korać, J. Vukojević, and A. Knežević, “Potential of pleurotus ostreatus mycelium for selenium absorption,” Scientific World Journal, vol. 2014, 2014, doi: 10.1155/2014/681834.
[39] M. Haneef, L. Ceseracciu, C. Canale, I. S. Bayer, J. A. Heredia-Guerrero, and A. Athanassiou, “Advanced Materials from Fungal Mycelium: Fabrication and Tuning of Physical Properties,” Sci Rep, vol. 7, no. December 2016, pp. 1–11, 2017, doi: 10.1038/srep41292.
Notes
*
Research article
Author notes
aCorresponding author. E-mail: adriana.suesca@uamerica.edu.co
Additional information
How to
cite this article: A. Suesca Díaz, W. S. Pedraza
Baquero, Y. Morales Hernández, “Evaluation of the Production of
a Biocomposite Material from a Lignocellulosic Waste
Mixture using Mycelium of Pleurotus
ostreatus” Ing. Univ. vol. 29, 2025. https://doi.org/10.11144/Javeriana.iued29.epbm
