




APA
ISO 690-2
Harvard
Haga clic en un formato de citación
Implicaciones en el tratamiento de pacientes con cáncer de mama y alteraciones en ARID1A
ARID1A Alterations and Their Clinical Significance in Breast Cancer
Luisana Molina Pimienta, Juan Camilo Salgado Sánchez, Íngrid Hernández
Implicaciones en el tratamiento de pacientes con cáncer de mama y alteraciones en ARID1A
Universitas Medica, vol. 64, núm. 1, 2023
Pontificia Universidad Javeriana
Luisana Molina Pimienta a luisana.molina@javeriana.edu.co
Pontificia Universidad Javeriana, Colombia
Juan Camilo Salgado Sánchez
Pontificia Universidad Javeriana, Colombia
Íngrid Hernández
Fundación Universitaria Juan N. Corpas, Colombia
Resumen: ARID1A (AT-rich interaction domain 1A) es una subunidad de los complejos SWI/SNF específicamente mutada en ~20% de los cánceres humanos primarios. La inactivación de ARID1A a través de mutaciones somáticas y otros mecanismos epigenéticos da como resultado la pérdida de las funciones de guardián y cuidador en las células, lo que promueve la iniciación del tumor. Se ha documentado una correlación entre mutaciones de pérdida de función en ARID1A y la presencia de mutaciones activadoras en PIK3CA, pérdida de la expresión de PTEN y la pérdida de la función de p53. Las mutaciones de ARID1A estaban presentes en el 2,5% de todos los cánceres de mama; no obstante, el porcentaje de cáncer de mama con mutaciones en ARID1A aumenta en los cánceres metastásicos un 12%, o en los inflamatorios, un 10%. La pérdida de la función de la ARID1A en cáncer de mama se adquiere con mayor frecuencia posterior al tratamiento y está asociada con la resistencia al tratamiento hormonal y con agentes quimioterapéuticos. Además, conduce a una reparación deficiente de las rupturas de doble cadena, que sensibilizan las células a los inhibidores de PARP. Por último, las alteraciones en ARID1A podrían ser un biomarcador de respuesta a inhibidores de punto de control.
Palabras clave:ARID1A, cáncer de mama, complejo SWI/SNF, agentes terapéuticos.
Abstract: ARID1A (AT-rich interaction domain 1A), is a subunit of the SWI/SNF complexes specifically mutated in ~20% of primary human cancers. Inactivation of ARID1A through somatic mutations and other epigenetic mechanisms results in loss of gatekeeper and caretaker functions in cells, promoting tumor initiation. A correlation has been documented between loss-of-function mutations in ARID1A and the presence of activating mutations in PIK3CA, loss of PTEN expression and loss of p53 function. ARID1A mutations were present in 2.5% of all breast cancers. However, the percentage of breast cancer with ARID1A mutations increases in metastatic 12% or inflammatory 10% cancers. Loss of ARID1A function in breast cancer is most frequently acquired post-treatment and is associated with resistance to hormonal treatment and chemotherapeutic agents. In addition, it leads to deficient repair of double-strand breaks, sensitizing cells to PARP inhibitors. Finally, alterations in ARID1A could be a biomarker of response to checkpoint inhibitors.
Keywords: ARID1A, breast cancer, SWI/SNF complex, therapeutic targets.
Introducción
El complejo de remodelación de la cromatina SWI/SNF altera la estructura del nucleosoma de una manera dependiente del trifosfato de adenosina, que afecta la accesibilidad de la cromatina y la expresión génica. Además, promueve la reparación de rupturas de la doble cadena (DSB), mediante el reclutamiento de proteínas reparadoras del ADN, tanto de la vía de reparación de unión de extremos no homólogos como en la recombinación homóloga (1).
En células somáticas, el complejo SWI/SNF se divide en varios complejos de factor asociado al gen 1 relacionado con Brahma (BAF), de acuerdo con sus funciones, y cada complejo consta de alrededor de 10 subunidades. Se han identificado al menos 29 subunidades como constituyentes de varios complejos SWI/SNF de mamíferos, lo que hace que tenga una variación notable (2).
La AT-rich interaction domain 1A (ARID1A) es una subunidad de los complejos SWI/SNF específicamente mutada en ~20% de los cánceres humanos primarios (3), también llamada BAF250a, p270 o SMARCF1, que contiene un dominio ARID de unión a ADN de 100 aminoácidos. La subfamilia ARID1 contiene dos miembros (ARID1A y ARID1B) que comparten aproximadamente un 80% de homología de aminoácidos dentro de sus dominios ARID y un 50% de homología en todas partes (4). Las subunidades del complejo BAF (incluidos ARID1A y SMARCA2) son necesarias para la unión de extremos no homólogos(5).
El ARID1A está ubicado en el cromosoma 1p36.11, se expresa ampliamente en el núcleo y varía de acuerdo con el ciclo celular, alcanzando su punto máximo durante G0/G1 (6). Su modificación postraducción, incluida la acetilación de N-16-lisina y la fosforilación de serina/treonina, puede regular la expresión de proteínas o las interacciones proteína-proteína(7).
En términos generales, los genes supresores de tumores se pueden clasificar en dos clases: cuidadores y guardianes (8). Los primeros controlan la proliferación celular, generalmente mediante la regulación del ciclo celular o la promoción de la apoptosis; mientras que los cuidadores mantienen la integridad del genoma. La inactivación de ARID1A a través de mutaciones somáticas y otros mecanismos epigenéticos da como resultado la pérdida de las funciones de guardián y cuidador en las células, lo que promueve la iniciación del tumor (9). La eliminación de ARID1A, pero no de ARID1B, anula la detención del ciclo celular asociada con la diferenciación en osteoblastos, lo que sugiere que la pérdida de ARID1A puede favorecer la formación de complejos que contienen ARID1B e impulsar la proliferación (10).
El ARID1A es el que muta con mayor frecuencia entre todos los genes que codifican subunidades de complejos SWI/SNF (11). Los linajes tumorales asociados con mutaciones de ARID1A son: carcinoma de células claras de ovario, carcinoma endometroide del uterino, cáncer gástrico, carcinoma hepatocelular, adenocarcinoma esofágico, cáncer de mama, cáncer de páncreas, carcinoma de vejiga de células transicionales, cáncer renal y colangiocarcinoma (12-14).
La mayoría de las mutaciones de ARID1A (>97%) dan como resultado una disminución de la expresión de las proteínas de la subunidad (15), mutaciones sin sentido o de cambio de marco de lectura detectadas en todo el gen (7). El análisis de hibridación genómica comparativa de los tumores de mama primario demostró la pérdida de material cromosómico que codifica una copia de ARID1A en el 35% de las muestras, sin encontrar ningún mutación en el alelo restante (16).
Evaluar las mutaciones conductoras específicas del cáncer de mama, permitirá conocer mejores estrategias para mejorar la eficacia clínica de los tratamientos actuales y nuevos. Por medio de la presente revisión se resume la evidencia actual sobre la relación entre las alteraciones del gen ARID1A y el tratamiento de pacientes con cáncer de mama. El objetivo es resumir la evidencia actual de la mutación conductora en el gen de la proteína ARID1A en los pacientes con cáncer de mama y sus posibles implicaciones en el tratamiento.
Métodos
Se buscó literatura sobre el tema, relativa a los últimos 20 años, en las bases de datos de Medline, Embase, Scopus, LILACS y Web of Science, utilizando los términos MeSH: Breast Carcinoma OR Breast Neoplasms OR Breast Cancer AND ARID1A AND Treatment response OR response OR treatment. Se incluyeron artículos generales, todos los diseños de estudios de ciencias básicas, estudios clínicos, cohortes y revisiones de la literatura, en idiomas inglés y español. Adicionalmente, se incluyeron recomendaciones de expertos y bibliografía de artículos.
Se utilizó la herramienta Rayyan-Intelligent Systematic Review para seleccionar los estudios obtenidos, a partir de la búsqueda por títulos y resumen. En una segunda fase se revisó el texto completo y los artículos con datos potencialmente relevantes se incluyeron para una revisión detallada.
Cáncer de mama
El cáncer de mama es uno de los cánceres más frecuentes en el mundo, tanto que en el 2020 se diagnosticaron 2,3 millones de mujeres con cáncer de mama y se registraron 685000 muertes (17). El 20%-30% de las pacientes con cáncer de mama pueden desarrollar metástasis después del diagnóstico, y aproximadamente el 90% de las muertes relacionadas con el cáncer se atribuyen a la metástasis (18). Desde la utilización de las técnicas de secuenciación de próxima generación se han descrito múltiples alteraciones genéticas relacionadas con el cáncer de mama, y algunos de los genes conductores previamente descritos son: PIK2CA, TP53, ERBB2, AKT1, BCRA, GATA3, CDH1, etc. (19). Las mutaciones de ARID1A estaban presentes en el 2,5% de todos los cánceres de mama (3). No obstante, el porcentaje de cáncer de mama con mutaciones en ARID1A aumenta en los cánceres metastásicos un 12%, o inflamatorios, un 10% (20), secundario a que la pérdida de la función de la ARID1A se adquiere con mayor frecuencia posterior al tratamiento. En el cáncer de mama metastásico, el ARID1A es la mutación más frecuente del complejo SWI/SNF (21).
Cáncer con deficiencia de ARID1A
La pérdida de ARID1A puede tener muchos efectos en los complejos SWI/SNF que resultan en disfunción transcripcional, incluida la alteración de la remodelación de nucleosomas, ensamblaje de las variantes del complejo SWI/SNF y orientación de loci genómicos específicos (7).
El ARID1A y el ARID1B se han caracterizado como miembros mutuamente exclusivos de los complejos SWI/SNF, variantes de BAF (22). En consecuencia, los tumores con deficiencia de ARID1A presentan una dependencia de ARID1B y exhiben una mayor vulnerabilidad a los inhibidores de las vías de reparación del ADN (23). Silenciar el ARID1B alteró la proliferación celular en células cancerosas con mutaciones ARID1A pero no en células con ARID1A de tipo salvaje (24). Por ende, el ARID1B puede ser un objetivo terapéutico prometedor de cánceres con mutación ARID1A.
Varios estudios han mostrado una correlación entre mutaciones de pérdida de función en ARID1A y la presencia de mutaciones activadoras en PIK3CA (25,26), pérdida de la expresión de PTEN (27) y pérdida de la función de p53 (28). No se descarta un proceso de cooperación oncogénica en la mutación de ARID1A y los genes de la vía PI3K.
Implicaciones de ARID1A en el cáncer de mama e implicaciones en el tratamiento
En el cáncer de mama, la expresión baja del ARNm de ARID1A, se ha asociado con características agresivas que incluyen un grado más alto, un estadio más alto, mayor Ki-67 y con el subtipo triple negativo (29). Mamo et al. (30) mostraron que la baja expresión de ARID1A en líneas celulares de cáncer de mama está asociada con un grado tumoral alto, inactivación de p53 y alta expresión de ERBB2. El principal mecanismo subyacente de la baja expresión de ARNm de ARID1A (86,4%) en este tipo de cáncer es la metilación del promotor, acompañada de una modificación de histonas (H3K27Me3) (31).
Takao et al. (32) evaluaron la expresión de ARID1A por inmunohistoquímica en muestras de tejido con cáncer de mama, y su pérdida parcial menor del 50% se correlacionó con una disminución de la supervivencia libre de enfermedad y con un aumento de la expresión de RAB11FIP1 (proteína 1 que interactúa con la familia RAB11), que ayuda a la progresión del cáncer de mama.
En el cáncer de mama con receptores hormonales positivos, el receptor de estrógenos (RE) es el principal impulsador de la proliferación celular, y las terapias endocrinas han tenido un éxito importante en la práctica clínica. No obstante, la mayoría de los cánceres desarrolla resistencia a dicha terapia, y solo en el 18% de los casos el mecanismo de resistencia está vinculado con el receptor propiamente dicho (33).
Hay discrepancias entre los resultados de los estudios que intentan explicar el peor pronóstico y la resistencia a la terapia endocrina en los cánceres de mama con pérdida de ARID1A (34). Xu et al. (35), utilizando la tecnología CRISPR CAS9, encontraron que células de cáncer de mama con ARID1A knockout tenían una mayor proliferación en condiciones de deprivación de estrógenos, en comparación con controles, y que estas células aumentaron significativamente la expresión de tipo basal como KRT6, KRT15, KRT5, CD44 y TP63; mientras que la expresión de marcadores luminales, como GATA3, ER, FOXA1, KRT8, TFF3, WISP2/CCN5 y CITED1, fueron regulados a la baja, considerando que ARID1A participa en el mantenimiento de la identidad luminal mediante la modulación de la plasticidad celular.
Nagarajan et al. (36), utilizando pantallas de CRISPR, documentaron que el agotamiento de ARID1A (es decir, el enriquecimiento de ARN guía dirigidos a ARID1A) se asoció con la resistencia a medicamentos como fulvestrant. Además, mediante ChIP-seq se evidenció que más del 78% de todos los eventos de unión de ARID1A se compartieron con el RE, FOXA1 o ambas, mostrando que ARID1A es parte del complejo RE. Además, al usar ARN seq, la deleción de ARID1A resultó en la inducción de los genes reprimidos, incluso en ausencia de un antagonista de RE. Por último, la eliminación de ARID1A resultó en la pérdida de la unión de histona desacetilasa 1 (HDAC) y en un aumento de la acetilación de lisina de histona 4.
Los cánceres de mama triplenegativos carecen de los receptores para estrógeno, progesterona y Her2/neu; aparte de ser tumores heterogéneos y agresivos (37). La quimioterapia basada en Paclitaxel es una de las estrategias para tratar este tipo de cáncer. Lin et al. (38) evaluaron en líneas celulares de cáncer de mama que la expresión de ARID1A se correlacionó negativamente con la concentración 50 de Paclitaxel en las muestras analizadas y una respuesta más deficiente basada en los análisis de Kaplan‐Meier.
En el 2015 se publicó un artículo que informó que los inhibidores de PARP (Poli ADP ribosa polimerasa), diseñados para el tratamiento de cánceres con deficiencia en la reparación de ADN por mutaciones de BRCA 1/2 u otros genes, disminuyeron selectivamente la supervivencia de las células con expresión disminuida de ARID1A y que proporcionaron una estrategia de tratamiento para los pacientes con cáncer de mama y mutación ARID1A (39). Esto, relacionado con que ARID1A es una de las muchas proteínas de unión a ATR y, por lo tanto, es una proteína reclutada por DSB, la pérdida de ARID1A conduce a una reparación deficiente DSB, sensibilizando a las células a los tratamientos que inducen dobles rupturas, como la radiación y los inhibidores de PARP (5).
Al igual que BRCA1, ARID1A interactúa físicamente con la topoisomerasa IIα, que le permite la separación de cromátidas hermanas recién replicadas. Ello garantiza una adecuada segregación durante la mitosis, y la pérdida de ARID1A podría contribuir a la formación de puentes durante la anafase que dan como resultado aneuploidía y poliploidía (40). Además, resistencia al alterar el efecto citotóxico del etopósido (41).
La vía PI3K/AKT/mTOR es importante para la transducción de señales en respuesta a estímulos extracelulares, al regular funciones como proliferación, crecimiento, supervivencia, motilidad y metabolismo (42). Informamos un estudio fase Ia de alpelisib que incluyó pacientes con cáncer de mama metastásico o localmente avanzado (n = 36; 26,9%) y demostró un perfil de seguridad tolerable, una tasa de respuesta general en el cáncer de mama del 4,3%, una tasa de beneficio clínico de 17,4% y en combinación con letrozol en pacientes con cáncer de mama con RE + la tasa de respuesta general del 19% con una tasa de beneficio clínico del 35%. Así mismo, incluyó pacientes con alteración de ARID1A (13,2%) (43). En la reunión anual de la Sociedad Estadounidense de Oncología Clínica de 2022 (resumen 1006) se presentó el análisis retrospectivo de los datos del estudio de fase III SOLAR-1, donde el beneficio clínico de la combinación de alpelisib y fulvestrant fue independiente de alteraciones genéticas adicionales, incluidas TP53, CCND1, MAP3K1 y ARID1A (44).
Es probable que la mutación de ARID1A provoque la situación de adicción al potenciador del homólogo Zeste 2 (EZH2). La principal función de EZH2 es la supresión de genes epigenéticos como un componente esencial del complejo represivo 2 de Polycomb (PRC2), y es una de las dos enzimas catalíticas esenciales para la metilación de la histona H3 lisina 27 (45). La inhibición de la metiltransferasa EZH2 provoca letalidad sintética en células de cáncer de ovario con mutación en ARID1A (46). No obstante, actualmente hay un estudio fase II de tumores sólidos avanzados en reclutamiento que incluyen pacientes con cáncer de mama y mutación de ARID1A utilizando tazemetostat, un inhibidor de EZH2 aprobado por la FDA para pacientes con linfoma no Hodgkin de células B y sarcomas epitelioides (47).
Estudios previos presentaron que la inhibición de HDAC6 suprime notablemente el crecimiento de tumores con mutación en ARID1A con efectos inmunomoduladores en el microambiente tumoral (48,49). Hay datos sobre el efecto sinérgico de los inhibidores de HDAC6 e inhibidores de punto de control en pacientes con cáncer de ovario (50), pero se requieren más estudios para evaluar efecto en cáncer de mama.
Por medio de análisis proteómico se identificó que una proteína clave del proceso de reparación del ADN por apareamiento erróneo (mismatch repair, MMR), MSH2, interacciona con ARID1A, y en un ensayo cuantitativo de MMR se identificó una capacidad disminuida de MMR en líneas celulares con ARID1A knockdown y expresión de proteína MMR intacta (MSH2, MLH1 y MSH6); además, encontramos que una expresión más baja de ARID1A se correlacionaba con una capacidad más baja de MMR. En pacientes con tumores sin alteración de MMR, la mutación ARID1A se asoció con un mejor pronóstico (39).
En una cohorte de 3403 pacientes con 9 tipos de cáncer, incluyendo cáncer de mama y una prevalencia de alteraciones de ARID1A >5%, la inestabilidad de microsatélites y la alta carga mutacional del tumor son significativamente más frecuentes en tumores con ARID1A mutados vs. ARID1A tipo salvaje (20% vs 0,9% [p<0,001]; y 26% vs. 8,4% [p<0,001], respectivamente). Además, la mediana de supervivencia libre de progresión después de inmunoterapia con bloqueo de punto de control fue más larga en pacientes con ARID1A mutado (11 meses frente a 4 meses, con p = 0,006) y en el análisis multivariado la mutación de ARID1A predice una supervivencia libre de progresión (recombinación homóloga con un intervalo de confianza del 95%: 0,61 [0,39-0,94]; p = 0,02), independiente de la inestabilidad de microsatélites o la carga mutacional (51).
Estos hallazgos podrían sugerir un papel predictivo para la terapia con inhibidores de punto de control. No obstante, las alteración de ARID1A y su correlación con infiltrados de linfocitos varían según el tipo de cáncer. En los casos de cáncer de endometrio, colon y estómago se ha encontrado un aumento de las infiltrados de células T CD8+, a diferencia del cáncer de células claras renales, que presenta un infiltrado bajo en este tipo de célula (52).
Las alteraciones en ARID1A pueden aumentar la expresión de PD-L1 por DSB. Los DSB pueden regular al alza la expresión de PD-L1 en las células tumorales, lo que requiere la vía de señalización ATM/ATR/Chk1 y CD274 (codifica PD-L1) es un gen diana ARID1A directo (53).
De acuerdo con lo anterior, las terapias dirigidas que se han estudiado ampliamente en tumores con alteración de ARID1A son inhibidores de PARP, vía PI3K/AKT/mTOR, ATR, EZH2 e histona desacetilasa 6. Además, de la inmunoterapia a través de los inhibidores de punto de control (figura 1).
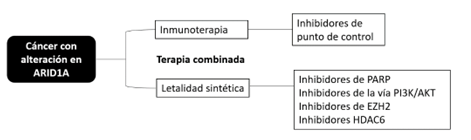
Conclusiones
El ARID1A es un gen supresor de tumores con función de guardián y cuidador en las células, que presenta alteraciones relacionadas con la disminución de su expresión en pacientes con cáncer de mama. La expresión disminuida por ARNm o por inmunohistoquímica está vinculada con características de mal pronóstico. Además, podría ser un biomarcador para escoger una terapia dirigida relacionada con letalidad sintética, inmunoterapia o combinada en pacientes con cáncer de mama y alteración de ARID1A.
Conflicto de intereses
Los autores no declaran conflictos de intereses.
Referencias
1. Tang L, Nogales E, Ciferri C. Structure and function of SWI/SNF chromatin remodeling complexes and mechanistic implications for transcription. Prog Biophys Mol Biol [internet]. 2010 Jun;102(2-3):122-8. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S0079610710000295
2. Harrod A, Lane KA, Downs JA. The role of the SWI/SNF chromatin remodelling complex in the response to DNA double strand breaks. DNA Repair (Amst) [internet]. 2020 Sep;93:102919. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S1568786420301671
3. Kadoch C, Hargreaves DC, Hodges C, Elias L, Ho L, Ranish J, et al. Proteomic and bioinformatic analysis of mammalian SWI/SNF complexes identifies extensive roles in human malignancy. Nat Genet [internet]. 2013 Jun 5;45(6):592-601. Disponible en: http://www.nature.com/articles/ng.2628
4. Wilsker D, Probst L, Wain H, Maltais L, Tucker P, Moran E. Nomenclature of the ARID family of DNA-binding proteins. Genomics [internet]. 2005 Aug;86(2):242-51. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S0888754305000923
5. Watanabe R, Ui A, Kanno S, Ogiwara H, Nagase T, Kohno T, et al. SWI/SNF factors required for cellular resistance to DNA damage include ARID1A and ARID1B and show interdependent protein stability. Cancer Res [internet]. 2014 May 1;74(9):2465-75. Disponible en: http://cancerres.aacrjournals.org/lookup/doi/10.1158/0008-5472.CAN-13-3608
6. Flores-Alcantar A, Gonzalez-Sandoval A, Escalante-Alcalde D, Lomelí H. Dynamics of expression of ARID1A and ARID1B subunits in mouse embryos and in cells during the cell cycle. Cell Tissue Res. 2011 Jul 7;345(1):137-48. https://doi.org/10.1007/s00441-011-1182-x
7. Wu JN, Roberts CWM. ARID1A mutations in cancer: another epigenetic tumor suppressor? Cancer Discov. 2013 Jan;3(1):35-43. https://doi.org/10.1158/2159-8290.CD-12-0361
8. Kinzler KW, Vogelstein B. Cancer-susceptibility genes: gatekeepers and caretakers. Nature [internet]. 1997 Apr 24;386(6627):761, 763. Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/9126728
9. Wu R-C, Wang T-L, Shih I-M. The emerging roles of ARID1A in tumor suppression. Cancer Biol Ther [internet]. 2014 Jun 11;15(6):655-64. Disponible en: http://www.tandfonline.com/doi/abs/10.4161/cbt.28411
10. Wilson BG, Roberts CWM. SWI/SNF nucleosome remodellers and cancer. Nat Rev Cancer [internet]. 2011 Jul 9;11(7):481-92. Disponible en: https://www.nature.com/articles/nrc3068
11. Shain AH, Pollack JR. The spectrum of SWI/SNF mutations, ubiquitous in human cancers. PLoS One. 2013 Jan 23;8(1):e55119. https://doi.org/10.1371/journal.pone.0055119
12. Wiegand KC, Lee AF, Al-Agha OM, Chow C, Kalloger SE, Scott DW, et al. Loss of BAF250a (ARID1A) is frequent in high-grade endometrial carcinomas. J Pathol. 2011 Jul;224(3):328-33. https://doi.org/10.1002/path.2911
13. Wiegand KC, Shah SP, Al-Agha OM, Zhao Y, Tse K, Zeng T, et al. ARID1A Mutations in endometriosis-associated ovarian carcinomas. N Engl J Med. 2010 Oct 14;363(16):1532-43. https://doi.org/10.1056/NEJMoa1008433
14. Jones S, Li M, Parsons DW, Zhang X, Wesseling J, Kristel P, et al. Somatic mutations in the chromatin remodeling gene ARID1A occur in several tumor types. Hum Mutat. 2012 Jan 23;33(1):100-3. https://doi.org/10.1002/humu.21633
15. St. Pierre R, Kadoch C. Mammalian SWI/SNF complexes in cancer: emerging therapeutic opportunities. Curr Opin Genet Dev [internet]. 2017 Feb;42:56-67. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S0959437X17300278
16. Cornen S, Adelaide J, Bertucci F, Finetti P, Guille A, Birnbaum DJ, et al. Mutations and deletions of ARID1A in breast tumors. Oncogene [internet]. 2012 Sep 20;31(38):4255-6. Disponible en: https://www.nature.com/articles/onc2011598
17. World Health Organization. World cancer day 2021: spotlight on IARC research related to breast cancer [internet]. [Citado 2021 dic 12]. Disponible en: https://www.iarc.who.int/featured-news/world-cancer-day-2021/
18. Liang Y, Zhang H, Song X, Yang Q. Metastatic heterogeneity of breast cancer: molecular mechanism and potential therapeutic targets. Semin Cancer Biol [internet]. 2020 Feb;60:14-27. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S1044579X1930063X
19. Fishbein L, Leshchiner I, Walter V, Danilova L, Robertson AG, Johnson AR, et al. Comprehensive molecular characterization of pheochromocytoma and paraganglioma. Cancer Cell [internet]. 2017 feb;31(2):181-93. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S1535610817300016
20. Yates LR, Knappskog S, Wedge D, Farmery JHR, González S, Martincorena I, et al. Genomic evolution of breast cancer metastasis and relapse. Cancer Cell [internet]. 2017 Aug;32(2):169-184.e7. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S1535610817302970
21. Cheng X, Zhao J-X, Dong F, Cao X-C. ARID1A mutation in metastatic breast cancer: a potential therapeutic target. Front Oncol. 2021 Nov 4;11. https://doi.org/10.3389/fonc.2021.759577/full
22. Martens JA, Winston F. Recent advances in understanding chromatin remodeling by Swi/Snf complexes. Curr Opin Genet Dev [internet]. 2003 Apr;13(2):136-42. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S0959437X03000224
23. Shen J, Peng Y, Wei L, Zhang W, Yang L, Lan L, et al. ARID1A Deficiency impairs the DNA damage checkpoint and sensitizes cells to PARP inhibitors. Cancer Discov. 2015 Jul;5(7):752-67. https://doi.org/10.1158/2159-8290.CD-14-0849
24. Simmons DJ, Parvin C, Smith KC, France P, Kazarian L. Effect of rotopositioning on the growth and maturation of mandibular bone in immobilized rhesus monkeys. Aviat Space Environ Med [internet]. 1986 Feb;57(2):157-61. Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/3954704
25. Samartzis EP, Gutsche K, Dedes KJ, Fink D, Stucki M, Imesch P. Loss of ARID1A expression sensitizes cancer cells to PI3K- and AKT-inhibition. Oncotarget. 2014 Jul 30;5(14):5295-303. https://doi.org/10.18632/oncotarget.2092
26. Yamamoto S, Tsuda H, Takano M, Tamai S, Matsubara O. PIK3CA mutations and loss of ARID1A protein expression are early events in the development of cystic ovarian clear cell adenocarcinoma. Virchows Arch. 2012 Jan 26;460(1):77-87. https://doi.org/10.1007/s00428-011-1169-8
27. Suryo Rahmanto Y, Shen W, Shi X, Chen X, Yu Y, Yu Z-C, et al. Inactivation of Arid1a in the endometrium is associated with endometrioid tumorigenesis through transcriptional reprogramming. Nat Commun [internet]. 2020 Dec 1;11(1):2717. Disponible en: http://www.nature.com/articles/s41467-020-16416-0
28. Allo G, Bernardini MQ, Wu R-C, Shih I-M, Kalloger S, Pollett A, et al. ARID1A loss correlates with mismatch repair deficiency and intact p53 expression in high-grade endometrial carcinomas. Mod Pathol [internet]. 2014 Feb 26;27(2):255-61. Disponible en: http://www.nature.com/articles/modpathol2013144
29. Zhang X, Zhang Y, Yang Y, Niu M, Sun S, Ji H, et al. Frequent low expression of chromatin remodeling gene ARID1A in breast cancer and its clinical significance. Cancer Epidemiol [internet]. 2012 Jun;36(3):288-93. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S1877782111001196
30. Mamo A, Cavallone L, Tuzmen S, Chabot C, Ferrario C, Hassan S, et al. An integrated genomic approach identifies ARID1A as a candidate tumor-suppressor gene in breast cancer. Oncogene [internet]. 2012 Apr 19;31(16):2090-100. Disponible en: https://www.nature.com/articles/onc2011386
31. Zhang X, Sun Q, Shan M, Niu M, Liu T, Xia B, et al. Promoter hypermethylation of ARID1A gene is responsible for its low mRNA expression in many invasive breast cancers. PLoS One. 2013;8(1):e53931. Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/23349767
32. Takao C, Morikawa A, Ohkubo H, Kito Y, Saigo C, Sakuratani T. Downregulation of ARID1A, a component of the SWI/SNF chromatin remodeling complex, in breast cancer. Cancer. 2017 Jan 1;8(1):1-8. https://doi.org/10.7150/jca.16602
33. Toy W, Shen Y, Won H, Green B, Sakr RA, Will M, et al. ESR1 ligand-binding domain mutations in hormone-resistant breast cancer. Nat Genet [internet]. 2013 Dec 3;45(12):1439-45. Disponible en: http://www.nature.com/articles/ng.2822
34. Farcas AM, Nagarajan S, Cosulich S, Carroll JS. Genome-wide estrogen receptor activity in breast cancer. Endocrinology. 2021 Feb 1;162(2). https://doi.org/10.1210/endocr/bqaa224/6025411
35. Xu G, Chhangawala S, Cocco E, Razavi P, Cai Y, Otto JE, et al. ARID1A determines luminal identity and therapeutic response in estrogen-receptor-positive breast cancer. Nat Genet [internet]. 2020 Feb 13;52(2):198-207. Disponible en: http://www.nature.com/articles/s41588-019-0554-0
36. Nagarajan S, Rao S V., Sutton J, Cheeseman D, Dunn S, Papachristou EK, et al. ARID1A influences HDAC1/BRD4 activity, intrinsic proliferative capacity and breast cancer treatment response. Nat Genet [internet]. 2020 Feb 1;52(2):187-97. Disponible en: https://www.nature.com/articles/s41588-019-0541-5
37. Denkert C, Liedtke C, Tutt A, von Minckwitz G. Molecular alterations in triple-negative breast cancer—the road to new treatment strategies. Lancet [internet]. 2017 Jun;389(10087):2430-42. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S0140673616324540
38. Lin Y-F, Tseng I-J, Kuo C-J, Lin H-Y, Chiu I-J, Chiu H-W. High-level expression of ARID1A predicts a favourable outcome in triple-negative breast cancer patients receiving paclitaxel-based chemotherapy. J Cell Mol Med [internet]. 2018;22(4):2458-68. Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/29392887
39. Shen J, Peng Y, Wei L, Zhang W, Yang L, Lan L, et al. ARID1A Deficiency Impairs the DNA Damage Checkpoint and Sensitizes Cells to PARP Inhibitors. Cancer Discov [internet]. 2015 Jul 1;5(7):752-67. Disponible en: https://aacrjournals.org/cancerdiscovery/article/5/7/752/5179/ARID1A-Deficiency-Impairs-the-DNA-Damage
40. Dykhuizen EC, Hargreaves DC, Miller EL, Cui K, Korshunov A, Kool M, et al. BAF complexes facilitate decatenation of DNA by topoisomerase IIα. Nature [internet]. 2013 May 22;497(7451):624-7. Disponible en: http://www.nature.com/articles/nature12146
41. Hu H-M, Zhao X, Kaushik S, Robillard L, Barthelet A, Lin KK, et al. A Quantitative Chemotherapy Genetic Interaction Map reveals factors associated with PARP inhibitor resistance. Cell Rep [internet]. 2018 Apr;23(3):918-29. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S2211124718304546
42. Engelman JA, Luo J, Cantley LC. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat Rev Genet [internet]. 2006 Aug;7(8):606-19. Disponible en: http://www.nature.com/articles/nrg1879
43. Juric D, Rodon J, Tabernero J, Janku F, Burris HA, Schellens JHM, et al. Phosphatidylinositol 3-kinase α-selective inhibition with alpelisib (BYL719) in PIK3CA -altered solid tumors: results from the first-in-human study. J Clin Oncol [internet]. 2018 May 1;36(13):1291-9. Disponible en: https://doi.org/10.1200/JCO.2017.72.7107
44. Juric D, Rugo HS, Reising A et. al. Alpelisib (ALT) + fulvestrant (FUL) in patients (pts) with hormone receptor-positive (HR+), human epidermal growth factor receptor 2-negative (HER2-) advanced breast cancer (ABC): Biomarker (BM) analyses by next-generation sequencing (NGS) from the SOLAR 1 study. J Clin Oncol. 2022;40(6 supl):1006. https://doi.org/10.1200/JCO.2022.40.16_suppl.1006
45. Yamagishi M, Uchimaru K. Targeting EZH2 in cancer therapy. Curr Opin Oncol [internet]. 2017 Sep;29(5):375-81. Disponible en: https://journals.lww.com/00001622-201709000-00011
46. Bitler BG, Aird KM, Garipov A, Li H, Amatangelo M, Kossenkov A V, et al. Synthetic lethality by targeting EZH2 methyltransferase activity in ARID1A-mutated cancers. Nat Med [internet]. 2015 Mar 16;21(3):231-8. Disponible en: http://www.nature.com/articles/nm.3799
47. Xu S, Tang C. The Role of ARID1A in tumors: tumor initiation or tumor suppression? Front Oncol [internet]. 2021;11:745187. Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/34671561
48. Bitler BG, Wu S, Park PH, Hai Y, Aird KM, Wang Y, et al. ARID1A-mutated ovarian cancers depend on HDAC6 activity. Nat Cell Biol [internet]. 2017 Aug 24;19(8):962-73. Disponible en: http://www.nature.com/articles/ncb3582
49. Liu Y, Li Y, Liu S, Adeegbe DO, Christensen CL, Quinn MM, et al. NK Cells Mediate synergistic antitumor effects of combined inhibition of HDAC6 and BET in a SCLC preclinical model. Cancer Res [internet]. 2018 Jul 1;78(13):3709-17. Disponible en: https://aacrjournals.org/cancerres/article/78/13/3709/625125/NK-Cells-Mediate-Synergistic-Antitumor-Effects-of
50. Fukumoto T, Fatkhutdinov N, Zundell JA, Tcyganov EN, Nacarelli T, Karakashev S, et al. HDAC6 inhibition synergizes with anti-PD-L1 therapy in ARID1A-inactivated ovarian cancer. Cancer Res [internet]. 2019 Nov 1;79(21):5482-9. Disponible en: https://aacrjournals.org/cancerres/article/79/21/5482/657614/HDAC6-Inhibition-Synergizes-with-Anti-PD-L1
51. Okamura R, Kato S, Lee S, Jimenez RE, Sicklick JK, Kurzrock R. ARID1A alterations function as a biomarker for longer progression-free survival after anti-PD-1/PD-L1 immunotherapy. J Immunother Cancer. 2020 Feb;8(1):e000438. https://doi.org//10.1136/jitc-2019-000438
52. Jiang T, Chen X, Su C, Ren S, Zhou C. Pan-cancer analysis of ARID1A alterations as biomarkers for immunotherapy outcomes. J Cancer [internet]. 2020;11(4):776-80. Disponible en: http://www.jcancer.org/v11p0776.htm
53. Wang L, Qu J, Zhou N, Hou H, Jiang M, Zhang X. Effect and biomarker of immune checkpoint blockade therapy for ARID1A deficiency cancers. Biomed Pharmacother [internet]. 2020 Oct;130:110626. Disponible en: https://linkinghub.elsevier.com/retrieve/pii/S0753332220308192
Notas de autor
a Autora de correspondencia: luisana.molina@javeriana.edu.co
Información adicional
Cómo citar: Molina Pimienta L, Salgado Sánchez JC, Hernández I. Implicaciones en el tratamiento de pacientes con cáncer de mama y alteraciones en ARID1A. Univ. Med. 2023;64(1). https://doi.org/10.11144/Javeriana.umed64-1.tpcm
